





После активации
В отличие от набухания ядер сперматозоидов, которое не зависит от активации ооцитов, все последующие ядерные трансформации сперматозоидов и ядерного материала ооцитов требуют активации ооцитов и, как следствие, снижения активности киназы p34 (cdc2) .
Прежде чем может начаться деконденсация, хромосомы как сперматозоидов, так и яйцеклетки должны конденсироваться и достичь «стадии хроматиновой массы». Весьма вероятно, что этот этап необходим для достижения синхронизации хроматина сперматозоидов и ооцитов перед деконденсацией.
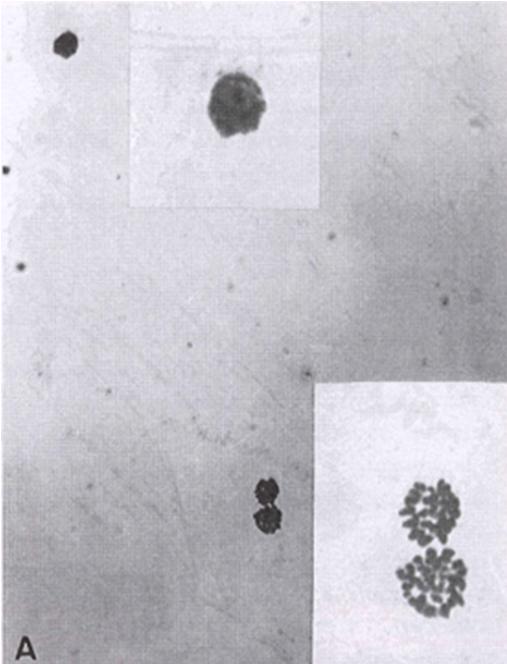
Ранняя телофаза примерно через 4,5 часа после инъекции. Обратите внимание, что на этом этапе ядро сперматозоида не деконденсируется, а остается набухшим. Также обратите внимание, что хромосомы ооцита и полярного тельца образуют две группы, неотличимые друг от друга.

Поздняя телофаза (также известная как стадия массы хроматина) вскоре после экструзии 1PB в однодневном ооците человека. Хроматин сперматозоидов остается набухшим, а хромосомы ооцитов плотно упакованы. На этом этапе хромосомы ооцитов и сперматозоидов синхронизируются перед деконденсацией.
Асинхронность трансформации хромосом сперматозоидов и ооцитов может приводить к образованию одного пронуклеуса. Такая асинхронность после ИКСИ может быть вызвана техническими ошибками инъекции, такими как чрезмерное количество PVP, введенного вместе со сперматозоидом. Поскольку факторы ооцитов, необходимые для образования пронуклеуса, существуют в цитоплазме только в течение относительно короткого периода времени, любые значительные препятствия для взаимодействия сперматозоидов и ооплазмы после инъекции сперматозоидов могут привести к тому, что в ядре сперматозоида не будет окна деконденсации и не будет образовываться пронуклеус.
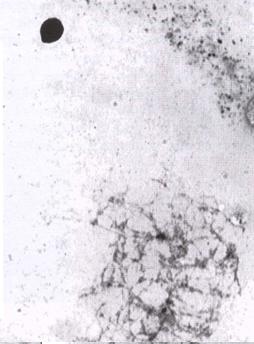
В этом человеческом ооците на следующий день после ИКСИ был обнаружен единственный пронуклеус. Головка сперматозоида опухла, что указывает на то, что она была введена правильно и вступила во взаимодействие с ооцитом, что привело к активации. Однако по неизвестным причинам он не деконденсировался. Полученный гаплоидный эмбрион будет нежизнеспособным.
Механизм оплодотворения после ИКСИ
В то время как глутатион отвечает за восстановление дисульфидных связей и его действие ограничивается независимым от активации ооцитов набуханием ядра сперматозоидов, SNDF требуется для последующей деконденсации ДНК (требует активации ооцитов), приводящей к образованию мужских пронуклеусов. Точная природа SNDF неизвестна, хотя было установлено, что она происходит из ядерного материала GV .

Определение сперматозоидов в образце яичек
Биопсия яичка, тонкоигольная аспирация и микродиссекция отдельных канальцев могут дать подвижные жизнеспособные сперматозоиды для ИКСИ. Идентификация жизнеспособных сперматозоидов в препарате гомогенизированных канальцев требует сортировки по множеству типов клеток, общих для канальцев яичка, чтобы найти наиболее подходящие сперматозоиды для инъекции. Сперматозоиды отбираются в первую очередь по подвижности, а во вторую – по морфологии. Сперматозоиды яичек незрелые, и их подвижность может проявляться в виде медленных или быстрых непрогрессирующих подергиваний, а иногда и в виде подвижности с прогрессированием вперед.
Идентифицировать сперматозоиды в препарате легче, когда плотность сперматозоидов, отражающая сперматогенез, в большей части ткани семенников нормальна; однако бывают случаи, когда сперматогенез не согласован по всем канальцам, и ткань демонстрирует мозаичный узор морфологии канальцев. В этом случае отдельные канальцы могут быть оценены во время микродиссекции, или ткань яичек может сопоставляться с помощью сеточного подхода, путем множественных аспираций тонкой иглой для определения местоположения отдельных очагов сперматогенеза. После этого можно исследовать конкретные области сперматогенеза. Отдельные канальцы, которые с большей вероятностью будут иметь участки, содержащие сперматозоиды, будут больше в диаметре и непрозрачны по сравнению с участками без активного сперматогенеза, и существует большая вероятность сперматогенеза в канальцах ближе к кровоснабжению.
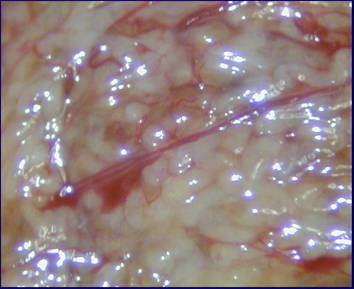
Крупным планом – семенные канальцы и кровоснабжение канальцев
Ткань яичек можно получить на месте рядом с лабораторией ЭКО или за ее пределами и доставить в лабораторию ЭКО. Кроме того, ткань может быть собрана в тот же день, что и пункция ооцитов, или перед пункцией ооцитов и, например , выдержана в культуре в течение ночи. Может быть криоконсервирована и сохранена в жидком азоте до тех пор, пока это не понадобится.
Канальцы яичка можно обрабатывать различными методами. Хотя это требует времени, отдельные канальцы можно раздавливать, когда канальцы визуализируются под микроскопом для препарирования. Для этого используется не режущий инструмент. Например, закругленный край стеклянной пипетки или предметное стекло сдвигается вниз и проводится по длине канальца, выталкивая содержимое канальца с одного конца. Этот процесс повторяется по мере необходимости. Объемные канальцы можно гомогенизировать, используя иглы и/или лезвия скальпеля, или используя стерильную ступку и гомогенизаторы ткани типа пестика. Гомогенат можно собрать в среде и центрифугировать, чтобы сконцентрировать содержимое канальцев для исследования. Ферментативное применение может повысить эффективность процедуры, увеличивая выход жизнеспособных сперматозоидов у лиц со сниженным сперматогенезом.
При активном сперматогенезе клетки в гомогенате будут представлены типами сперматозоидов и других клеток. Причем наиболее легко идентифицируемыми типами клеток являются зрелые сперматозоиды и красные кровяные тельца, которые загрязняют препарат. Движение может исходить от самого сперматозоида или косвенно от столкновения или отталкивания сперматозоидов от других клеток, или движения более крупных скоплений клеток от незрелых сперматозоидов, пока они еще прикреплены к клеткам. Это также может быть связано с наличием клеток из vasa efferentia или эпителиальных клеток проксимального отдела головки (S. Silber, не опубликовано). Эти клетки имеют реснички, которые выводят сперматозоиды из яичка в придаток яичка. Из-за неполного разделения сперматид это могут быть многохвостые сперматозоиды в ткани канальцев. Подводя подозреваемые многохвостые сперматозоиды к дну чашки с помощью микропипетки, можно распутать их хвосты. Морфология незрелых сперматозоидов может заметно отличаться от клеток в эякуляте. В частности, проксимальные капли могут быть более выраженными, поскольку остаточная цитоплазма еще не уменьшилась. Кроме того, могут быть обнаружены удлиненные сперматиды с полным образованием хвоста, но у которых головка сперматозоида все еще заключена в цитоплазму. И если нет зрелых сперматозоидов, можно рассмотреть эти сперматиды.
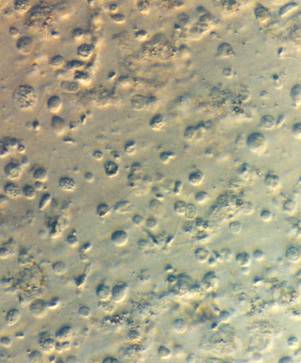
Нормальный сперматогенез в образце яичек
Ошибочная идентификация клеток как подвижных сперматозоидов в образце яичек
Иногда при первоначальном наблюдении в операционной отмечается подвижная сперма. Однако тщательное обследование в репродуктивной лаборатории с помощью более мощного инвертированного микроскопа позволяет сделать более точное определение. В этом случае «движущаяся клетка», изначально ошибочно идентифицированная как подвижный сперматозоид, впоследствии была правильно идентифицирована Dr. Silber как vasa efferentia или проксимальная эпителиальная клетка головки .